|
ИНФОРМАЦИЯ НЕОБХОДИМАЯ ДЛЯ ФОРМИРОВАНИЯ УПРАВЛЕНИЯ В СИСТЕМЕ ДЫХАНИЯ
[ information for control signals forming in respiration system ] Информация необходимая для формирования управления в системе дыхания - это информация о целях системы, информация о состоянии системы и её элементов, информация о среде системы, информация о результатах управления в прошлом. На основе этой информации управляющее звено системы, дыхательный центр формирует сигналы управления, которые с упреждением (прогнозирование, Трифонов Е.В., 1978) подаются к объекту управления системы - аппарату внешнего дыхания.
В любых системах организма, в том числе и в системе дыхания, входные сигналы могут восприниматься специализированными структурами - сенсорными системами. Рецепторы сенсорных систем (сенсорные рецепторы) являются их сенсорными входами.
Управление системой дыхания не может осуществляться без наличия информации о целях системы дыхания.
Главной целью (назначением) системы дыхания является осуществление эффективного внешнего дыхания - доставки из воздуха в организм кислорода и выведения газообразных конечных продуктов метаболизма (главным образом двуокиси углерода). Эффективное дыхание обеспечивается такой организацией взаимодействия структур и функций, которое могло бы минимальными структурами и минимальными действиями обеспечить достижение целей системы. Цели системы формируются и достигаются в соответствии с потенциальными и актуальными потребностями организма. Эти цели могут быть предопределены генетически, сформированы регулятором системы в процессе деятельности или назначены извне.
Информация об объекте управления - это информация о том, в каком состоянии (вдоха или выдоха) находится аппарат внешнего дыхания. Отсутствие этой информации лишает возможности управляющее звено (дыхательный центр) сформировать управление.
Более ста лет тому назад было обнаружено, что растяжение лёгких тормозит вдох, а спадение лёгких стимулирует его. Позже было установлено, что в лёгких существует два типа механорецепторов. Они отличаются друг от друга, во-первых, скоростью, с которой они адаптируются к продолжительному растяжению лёгких и, во-вторых, величиной порога чувствительности. Эти рецепторы были названы «быстроадаптирующимися рецепторами» и «медленноадаптирующимися рецепторами» растяжения лёгких. Медленноадаптирующиеся рецепторы реагируют на незначительные степени растяжения лёгких. При этом наступает отчетливое торможение вдоха. Эти рецепторы чувствительнее других, быстроадаптирующихся. Быстроадаптирующиеся рецепторы активируются при растяжении лёгких вдвое большем, чем объём вдоха или при быстром спадении лёгких. Их чувствительность ниже, а активность продолжается менее одной секунды.
Кроме этих двух типов рецепторов были обнаружены другие рецепторы, воспринимающие информацию о состоянии объекта управления. Эти рецепторы могут быть расположены субэпителиально, в бронхиальных гладких мышцах, в лёгочных кровеносных сосудах, в альвеолярных мембранах, в плевре, в перихондриальных участках. При этом главное значение имеют первые два типа рецепторов описанные выше. Информация от этих рецепторов по афферентным волокнам блуждающего нерва поступает в ядра блуждающего нерва. Ядра блуждающего нерва имеют хорошие связи с респираторными нейронами дыхательного центра ствола головного мозга.
На основании информации от медленноадаптирующихся рецепторов дыхательный центр формирует сигналы, тормозящие вдох, а на основании информации от быстроадаптирующихся рецепторов - сигналы, активирующие вдох.
Помимо указанных рецепторов и образуемых ими рефлексогенных зон определенную роль в восприятии информации о состоянии объекта управления играют рецепторы дыхательных мышц и суставов грудной клетки.
Для формирования управления дыхательному центру необходима также информация о результатах предшествующего управления. Основными управляемыми переменными системы дыхания (в соответствии с основными целями системы) являются напряжение двуокиси углерода, PaCO2 и напряжение кислорода в артериальной крови, PaO2 . Информация об этих переменных, получаемая структурами управляющего звена в момент времени t , используется для формирования управления в момент времени t+1. Известно, что с этими двумя управляемыми переменными связана еще одна управляемая переменная - концентрация водородных ионов в артериальной крови [H+] (показателем щёлочности крови является водородный показатель, pH). Значения этих главных переменных могут восприниматься периферическими рецепторами, поступать в соответствующие этим рецепторам нервные центры сенсорных систем, а от них - к взаимодействующему с ними дыхательному центру. В этом случае говорят о периферических нейрогенных механизмах влияния этих переменных на дыхательный центр. Помимо того, изменения концентрации водородных ионов, напряжения двуокиси углерода и напряжения кислорода могут восприниматься непосредственно специализированными нейронами дыхательного центра (хемочувствительные нейроны). В этом случае говорят о непосредственных центральных химических влияниях этих переменных.
Рассмотрим подробнее периферические и центральные механизмы получения информации об основных управляемых переменных системы дыхания.
Известно, что электрическая стимуляция каротидных и аортальных хеморецепторов, моделирующая реальные увеличение PaO2 и снижение PaCO2 , может вызвать следующие эффекты: снижение уровня напряжения кислорода в крови, PaO2, увеличение концентрации ионов водорода в крови, [H+] , увеличение уровня напряжения двуокиси углерода в артериальной крови, PaCO2 , повышение температуры тела (гипертермию), связанную с изменением щёлочно-кислотного равновесия, снижение артериального давления крови (гипотензия) и ухудшение кровоснабжения (хеморецепторная ишемия). Последние три эффекта реализуются не непосредственно системой дыхания, а опосредованно, взаимодействующей с ней системой кровообращения. Такое взаимодействие обусловлено наличием у данных систем общей цели. Информация о напряжении двуокиси углерода, концентрации водородных ионов и о напряжении кислорода от хеморецепторов дуги аорты и ствола аорты проходит по афферентным волокнам блуждающего нерва (X пара черепных нервов) к соответствующим ядрам. Аналогичная информация от хеморецепторов каротидного синуса проходит по афферентным волокнам языкоглоточного нерва (IX пара черепных нервов) к соответствующим ядрам продолговатого мозга. Ядра блуждающего и языкоглоточного нервов имеют либо непосредственные связи с дыхательным центром, либо опосредованные связи через ретикулярную формацию. В свою очередь ретикулярная формация может оказывать либо активирующие, либо тормозные влияния на дыхательный центр. В зависимости от характера информации, воспринимаемой периферическими хеморецепторами, дыхательный центр может либо усиливать лёгочную вентиляцию, либо снижать её. Не исключается возможность участия других хеморецепторов организма, воспринимающих информацию об управляемых переменных системы дыхания.
Периферические нейрогенные механизмы влияния данных переменных, из которых наиболее важными являются PaCO2 и [H+] , имеют меньшую значимость по сравнению с их центральным непосредственным воздействием на дыхательный центр.
|
Схема. Локализация ростральных (R) и каудальных (C) групп хемочувствительных нейронов (проекция на вентральную поверхность продолговатого мозга). Римскими цифрами обозначены корешки черепных нервов.
|
 |
|
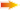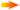 Дыхательный центр содержит несколько видов нейронов специализированных для восприятия концентрации водородных ионов, [H+], напряжения двуокиси углерода, PaCO2 , напряжения кислорода, PaO2 и, возможно, концентрации многих других веществ артериальной крови, питающей головной мозг. Это хемочувствительные нейроны. Они расположены в тонком слое ткани мозга у вентральной поверхности продолговатого мозга латеральнее пирамид. Выделяют две главные группы хемочувствительных нейронов: ростральная группа и каудальная группа. Кроме того, хемочувствительные нейроны обнаружены около ядер одиночного тракта, в области голубоватого места ромбовидной ямки и в гипоталамусе. Дыхательный центр содержит несколько видов нейронов специализированных для восприятия концентрации водородных ионов, [H+], напряжения двуокиси углерода, PaCO2 , напряжения кислорода, PaO2 и, возможно, концентрации многих других веществ артериальной крови, питающей головной мозг. Это хемочувствительные нейроны. Они расположены в тонком слое ткани мозга у вентральной поверхности продолговатого мозга латеральнее пирамид. Выделяют две главные группы хемочувствительных нейронов: ростральная группа и каудальная группа. Кроме того, хемочувствительные нейроны обнаружены около ядер одиночного тракта, в области голубоватого места ромбовидной ямки и в гипоталамусе.
Одни хемочувствительные нейроны расположены достаточно далеко от поверхности мозга, омываемой спинномозговой жидкостью, за гематоэнцефалическим барьером к прохождению веществ из крови. Так что любые изменения состава крови, отделенной гематоэнцефалическим барьером от спинномозговой и интерстициальной жидкости ткани мозга, не оказывают заметного влияния на эти хемочувствительные нейроны. Единственным веществом, для которого практически не существует барьеров для диффузии, является двуокись углерода, CO2 . Она может свободно диффундировать через любые ткани и гематоэнцефалический барьер вглубь ткани мозга. Полагают, что описанные хемочувствтительные нейроны, расположенные за гематоэнцефалическим барьером, могут реагировать либо непосредственно на увеличение напряжения двуокиси углерода, PaCO2 , либо на связанное с этим увеличение концентрации водородных ионов, [H+] . Двуокись углерода, беспрепятственно диффундирующая в интерстициальную жидкость ткани мозга, в спинномозговую жидкость, гидратируется с образованием угольной кислоты. Угольная кислота диссоциирует на бикарбонатный ион и ион водорода. В результате концентрация водородных ионов в интерстициальной ткани увеличивается пропорционально увеличению напряжения двуокиси углерода. Хемочувствительные нейроны могут реагировать именно на это увеличение концентрации водородных ионов. Результатом является увеличение лёгочной вентиляции.
Другие хемочувствительные нейроны расположены ближе к поверхности мозга и не защищены гематоэнцефалическим барьером. К ним могут свободно диффундировать многие вещества и в частности ионы водорода. Эти рецепторы высокочувствительны к изменению pH циркулирующей артериальной крови. В частности, они реагируют на ацидемию при мышечной работе и на метаболический ацидоз в артериальной крови, которой снабжается головной мозг. Наличие незащищенных гематоэнцефалическим барьером нейронов, чувствительных к концентрации водородных ионов, устраняет препятствие, связанное с наличием гематоэнцефалического барьера, практически непроницаемого для таких ионов как [H+] , [Cl–] или [HCO3–] . Если бы все хемочувствительные нейроны были защищены гематоэнцефалическим барьером, то это существенно снизило бы быстродействие регуляторов системы дыхания и влияние алкалоза или ацидоза могло бы проявиться значительно позже. Таким образом, наличие двух дублирующих структур для восприятия информации о результатах управления: хемочувствительных нейронов с большим быстродействием (незащищенных гематоэнцефалическим барьером) и хемочувствительных нейронов с низким быстродействием (защищенных гематоэнцефалическим барьером) обеспечивает высокую надёжность адаптации организма к изменениям состава жидкостей организма, к нарушениям кислотно-щёлочного равновесия.
Кроме хемочувствительных нейронов, специализированных на восприятии концентрации ионов водорода и напряжения двуокиси углерода, среди нейронов дыхательного центра обнаружили нейроны, чувствительные к напряжению кислорода. Такие хемочувствительные нейроны распределены в предкомплексе Бётцингера (Pre-Bötzinger complex), см. Литература. Иллюстрации, п. 11, 12, 13, 14, статью функциональные группы дыхательных нейронов ствола головного мозга. Вероятно, существуют хемочувствительные нейроны, специализированные для восприятия концентрации многих других веществ, влияющих на дыхание.
Для управления дыханием дыхательному центру необходима информация о среде. Такая информация поступает от взаимодействующих систем. Наиболее тесно с системой дыхания взаимодействует система кровообращения.
Известно, что помимо описанных выше хеморецепторов, в дуге и стволе аорты, в каротидном синусе расположены барорецепторы. Барорецепторы воспринимают информацию о давлении крови. Эта информация используется не только системой кровообращения, но и системой дыхания. В частности, увеличение давления крови может вызвать тормозные эффекты как со стороны системы кровообращения (вазодилатация), так и со стороны системы дыхания (уменьшение лёгочной вентиляции). Нервные волокна, которые заканчиваются барорецепторами, проходят в составе блуждающего и языкоглоточного нервов. Ядра этих нервов непосредственно или через ретикулярную формацию связаны с дыхательным центром, использующим информацию от взаимодействующих систем для организации структуры и функций системы дыхания. Помимо описанных выше специфических рефлексогенных зон, по всему кровеносному руслу распределены другие интерорецепторы, которые могут воспринимать информацию о химическом составе крови (хеморецепторы) и о физических переменных (механорецепторы) системы кровообращения.
Можно привести много других примеров поступления информации от взаимодействующих систем. Афферентные сигналы от мышц во время физических нагрузок, афферентные сигналы от сердца, от лёгочных сосудов малого круга кровообращения. В свою очередь, эмоциональные воздействия, сильные зрительные, тактильные, слуховые, болевые сигналы, сигналы о раздражении носовых и глоточных окончаний волокон тройничных нервов (V пара черепных нервов) сильными запахами - все эти сигналы можно рассматривать как информацию о среде организма в целом.
См.: Система дыхания: Cловарь,
Система дыхания: Литература. Иллюстрации,
Управление в системе дыхания: Литература. Иллюстрации,
Показатели деятельности системы дыхания.
|
«Я У Ч Е Н Ы Й И Л И . . . Н Е Д О У Ч К А ?»
Т Е С Т В А Ш Е Г О И Н Т Е Л Л Е К Т А
Предпосылка:
Эффективность развития любой отрасли знаний определяется степенью соответствия методологии познания - познаваемой сущности.
Реальность:
Живые структуры от биохимического и субклеточного уровня, до целого организма являются вероятностными структурами. Функции вероятностных структур являются вероятностными функциями.
Необходимое условие:
Эффективное исследование вероятностных структур и функций должно основываться на вероятностной методологии (Трифонов Е.В., 1978,..., ..., 2015, …).
Критерий: Степень развития морфологии, физиологии, психологии человека и медицины, объём индивидуальных и социальных знаний в этих областях определяется степенью использования вероятностной методологии.
Актуальные знания: В соответствии с предпосылкой, реальностью, необходимым условием и критерием...
...
о ц е н и т е с а м о с т о я т е л ь н о:
— с т е п е н ь р а з в и т и я с о в р е м е н н о й н а у к и,
— о б ъ е м В а ш и х з н а н и й и
— В а ш и н т е л л е к т !
|
♥ Ошибка? Щелкни здесь и исправь ее! Поиск на сайте E-mail автора (author): tryphonov@yandex.ru
|
