|
ГИПОФИЗ: ЗАДНЯЯ ДОЛЯ
[ neurohypophysis ]
ЭНДОКРИНОЛОГИЯ: ОГЛАВЛЕНИЕ = ENDOCRINOLOGY: CONTENTS
1. ОБЩАЯ ЭНДОКРИНОЛОГИЯ = GENERAL ENDOCRINOLOGY.
2. ЧАСТНАЯ ЭНДОКРИНОЛОГИЯ. НОРМА = SPECIAL ENDOCRINOLOGY. NORM.
2.1. Гипоталамус = The hypothalamus.
2.2. Гипофиз = The pituitary gland.
2.2.1. Макроструктура аденогипофиза = Macrostructure of adenohypophysis.
2.2.2. Микроструктура аденогипофиза = Microstructure of adenohypophysis.
2.2.3. Биоактивные вещества аденогипофиза = Bioactive substances of adenohypophysis.
2.2.4. Действие биоактивных веществ аденогипофиза = Actions of bioactive substances of adenohypophysis.
2.2.5. Макроструктура нейрогипофиза = Macrostructure of neurohypophysis.
2.2.6. Микроструктура нейрогипофиза = Microstructure of neurohypophysis.
2.2.7. Биоактивные вещества нейрогипофиза = Bioactive substances of neurohypophysis.
2.2.8. Действие биоактивных веществ нейрогипофиза = Actions of bioactive substances of neurohypophysis.
2.3. Щитовидная железа = The thyroid.
2.4. Околощитовидная железа = The parathyroid gland.
2.5. Hадпочечник = The adrenal cortex.
2.6. Островковый аппарат поджелудочной железы = Islet apparatus of the pancreas.
2.7. Половые железы = Gonads.
3. ЧАСТНАЯ ЭНДОКРИНОЛОГИЯ. ПАТОЛОГИЯ = SPECIAL ENDOCRINOLOGY. PATHOLOGY.
В РАЗРАБОТКЕ = UNDER CONSTRUCTION
Gray H., (1821–1865), Standring S., Ed. Gray's Anatomy: The Anatomical Basis of Clinical Practice = Г. Грей: Анатомические основы клинической практики. 39th ed., Churchill Livingstone, 2008, 1600 p.
Прекрасно иллюстрированное классическое учебное пособие и руководство, обновленное и дополненное коллективом современных авторов. В формате .chm. .
Доступ к данному источнику = Access to the reference.
URL: http://www.tryphonov.ru/tryphonov/serv_r.htm#0 quotation
Figure 21.13 The main systems controlling the endocrine secretory activities of the pituitary gland.
hypoph1_2.tif
hypoph2_1.tif
hypoph3_1.tif
hypoph4_1.tif = Figure 21.12 Median section through the hypophysis cerebri.
hypoth11_1.tif = Figure 21.16 A summary of the vasculature of the hypothalamic median eminence, infundibulum and the other regimes of the hypophysis cerebri
См. повторения рис. файл гипоталамус
Gardner D.G., Shoback D.M., Eds. Greenspan's Basic & Clinical Endocrinology. 9th ed., Lange, 2011, 960 p.
Table 4–5 Metabolic Effects of GH and IGF-1 In Vivo.
Table 4–6 Main Biologic Effect of the GH-IGF-1 Axis.
Table 4–7 Factors Affecting GH Secretion.
Table 4–9 Endocrine Tests of Hypothalamic-Pituitary Function.
В гипофизе выделяют две доли: передняя доля гипофиза (аденогипофиз) и задняя доля гипофиза (нейрогипофиз).
1. Передняя доля гипофиза. Переднюю долю гипофиза разделяют на три части: (а) передняя часть аденогипофиза, (б) промежуточная часть аденогипофиза, (в) туберальная часть аденогипофиза.
(а) Передняя часть аденогипофиза образована скоплениями эпителиальных клеток, образующих разветвленные тяжи - трабекулы, а также нерегулярные фолликулы. В промежутках между скоплениями клеток расположены синусоидальные кровеносные капилляры. Тонкие стенки синусоидов образованы рыхлой волокнистой соединительной тканью. Трабекулы и фолликулы содержат железистые клетки - эндокриноциты.
Различают три типа эндокриноцитов трабекул. Эндокриноциты, располагающиеся по периферии клеточных скоплений, содержат в своей цитоплазме секреторные гранулы, которые легко окрашиваются. В связи с этим такие клетки именуются хромофильными эндокриноцитами. Эндокриноциты, занимающие середину клеточного скопления, имеют нечеткие границы, поскольку их цитоплазма слабо окрашивается. Такие клетки именуются хромофобными эндокриноцитами.
Секреторные гранулы хромофильных эндокриноцитов могут окрашиваться либо основными, либо кислыми красителями. В соответствии с этим хромофильные эндокриноциты разделяют на базофильные и ацидофильные. Гранулы эндокриноцитов содержат гликопротеиды, являющиеся материалом для биосинтеза гормонов. Сравнительно крупные базофильные эндокриноциты в норме составляют ~4 ÷ 10 % общего количества аденоцитов. Выделяют две разновидности базофильных эндокриноцитов.
Клетки первой разновидности характеризуются округлой или овальной формой и эксцентрическим положением ядер. Их секреторные гранулы имеют диаметр ~200 ÷ 300 нм. Количество базофильных эндокриноцитов увеличивается при интенсивной выработке гонадотропных гормонов (гонадотропинов). В связи с этим такие клетки называются гонадотропоцитами, или гонадотропными эндокриноцитами. Одни из гонадотропоцитов вырабатывают фолликулостимулирующий гормон (фоллитропин), а другие - лютеинизирующий гормон (лютропин). Фоллитропин является средством управления формированием половых клеток. Лютропин является стимулирующим средством управления образованим жёлтого тела в яичнике, а также средством управления выработкой мужского полового гормона интерстициальными клетками яичка.
Базофильные клетки второй разновидности отличаются неправильной формой. Их секреторные гранулы очень мелкие (диаметр ~80 ÷ 150 нм) и легко окрашиваются альдегид-фуксином. Эти гранулы содержат меньше гликопротеинов, чем гранулы гонадотропоцитов. Клетки второй разновидности вырабатывают тиротропный гормон - тиротропин. Тиротропин является стимулирующим средством управления функциями фолликулярных эндокриноцитов щитовидной железы. Отсюда, клетки секретирующие тиротропин называются тиротропоцитами, или тиротропными эндокриноцитами.
Ацидофильные эндокриноциты содержат крупные плотные белковые гранулы, которые легко окрашиваются кислыми красителями. По размерам эти клетки несколько меньше базофильных эндокриноцитов. Их количество составляет ~30 ÷ 35 % всех аденоцитов передней доли гипофиза. Они имеют округлую или овальную форму. Ядра базофильных эндокриноцитов располагаются в центре клетки. У них хорошо развита гранулярная эндоплазматическая сеть. Ацидофильные эндокриноциты также имеют две разновидности. Первая разновидность ацидофильных эндокриноцитов - соматотропоциты, или соматотропные эндокриноциты. Одни соматотропоциты вырабатывают гормон роста, или соматотропин. Соматотропин является средством управления ростом организма. Другие соматотропоциты вырабатывают лактотропный гормон, или пролактин и потому их называют соответственно маммотропоцитами, или маммотропными эндокриноцитами и пролактиноцитами, или пролактотропными эндокриноцитами. Лактотропный гормон (пролактин) является средством управления биосинтезом молока в молочной железе. Интенсивность секреции пролактина усиливается у рожени́ц после родов, во время лактации и вскармливания новорожденного. Кроме того, пролактин продлевает существование и функции желтого тела в яичнике. Секреторные гранулы соматотропоцитов имеют шаровидную форму. Их диаметр составляет ~350 ÷ 400 нм. Маммотропоциты отличаются еще более крупными гранулами овальной или удлиненной формы (длина ~500 ÷ 600 нм, ширина ~100 ÷ 120 нм).
Хромофильные эндокриноциты включают ещё одну группу клеток, расположенных главным образом в центральной части передней доли гипофиза. Эти клетки называют кортикотропными эндокриноцитами, или кортикотропоцитами. Они вырабатывают адренокортикотропный гормон (АКТГ, или кортикотропин). Адренокортикотропный гормон является стимулирующим средством управления секрецией гормонов клетками пучковой зоны коркового вещества надпочечников. Кортикотропоциты имеют неправильную форму, дольчатые ядра, хорошо развитую эндоплазматическую сеть. Их секреторные гранулы имеют строение пузырьков, содержащих в своей полости плотную белковую сердцевину. Между мембраной пузырька и его сердцевиной остается светлое пространство.
Хромофобные клетки составляют ~60 % общего числа аденоцитов. Цитоплазма хромофобных эндокриноцитов слабо окрашивается. В ней не видны секреторные гранулы. Хромофобные клетки имеют разную степень развития и выполняют различные функции. Среди них существуют клетки, уже начавшие специализироваться в базофильные или ацидофильные клетки, но еще не успевшие накопить специфические секреторные гранулы. Другие же, хромофобные клетки, наоборот, являются вполне специализированными, но лишившимися своих базофильных или ацидофильных секреторных гранул вследствие интенсивной или длительной секреции. Небольшое количество неспециализированных хромофобных клеток относят к резервным клеткам.
Среди хромофобных клеток встречаются звездчатые (фолликулозвездчатые) клетки. Это клетки небольшого размера, имеющие длинные ветвистые отростки, посредством которых они соединяются в широкопетлистую сеть. Некоторые из отростков проходят между соседними эндокриноцитами и заканчиваются на стенках синусоидных капилляров. Иногда звездчатые клетки группируются в небольшие фолликулы. В полостях этих фолликулов накапливается гликопротеидный секрет. На апикальных поверхностях таких звездчатых клеток (со стороны просвета фолликула) имеются микроворсинки.
Функции аденогипофиза управляются посредством пептидов и аминов, секретируемых нейронами гипоталамуса. Эти нейросекреторные клетки расположены главным образом в медиальной зоне гипоталамуса, в его дугообразном ядре, в медиальной парвоцеллюлярной части паравентрикулярного и перивентрикулярного ядер.
(б) Средняя (промежуточная) часть аденогипофиза представляет собой узкую полоску эпителия. Эндокриноциты средней части способны вырабатывать белковый или слизистый секрет, который, накапливаясь между соседними клетками, приводит к формированию в средней части аденогипофиза фолликулоподобных кист. От задней части аденогипофиза эпителий средней части отделяется тонкой прослойкой рыхлой соединительной ткани.
Эндокриноциты средней части аденогипофиза вырабатывают меланоцитостимулирующий гормон (меланоцитотропин), а также липотропин - гормон, являющийся средством управления метаболизмом липидов.
(в) Туберальная часть аденогипофиза прилежит к ножке гипофиза и соприкасается с нижней поверхностью медианного возвышения гипоталамуса. Туберальная часть образована эпителиальными тяжами, состоящими из кубических клеток с умеренно базофильной цитоплазмой. От туберальных тяжей в переднюю часть аденогипофиза, так же как от эпителия средней части, отходят трабекулы. В некоторых клетках туберальных тяжей встречаются базофильные гранулы.
Кровоснабжение гипоталамуса и гипофиза выполняет функцию реализации управления аденогипофизарного гормонопоэза.
2. Задняя доля гипофиза (нейрогипофиз). Задняя доля гипофиза содержит в основном клетки эпендимы. Они имеют отростчатую или веретеновидную форму и называются питуицитами задней доли гипофиза. Их многочисленные тонкие отростки заканчиваются в адвентиции кровеносных сосудов или на базальной мембране гемакапилляров.
В задней доле гипофиза накапливаются антидиуретический гормон (вазопрессин) и окситоцин. Гормоны вазопрессин и окситоцин вырабатываются крупными пептидохолинергическими нейросекреторными клетками переднего гипоталамуса. Вазопрессин является средством управления реабсорбцией в мочевых канальцах почки. Окситоцин является средством управления сокращением мускулатуры матки. Аксоны нейросекреторных клеток переднего гипоталамуса собираются в гипоталамо-нейрогипофизарные пучки, входят в заднюю долю гипофиза, где заканчиваются крупными терминалями (тельца Херринга, или накопительные тельца), на стенках гемакапилляров.
Percy Theodore Herring (1872-1967), физиолог, врач, Великобритания.
|
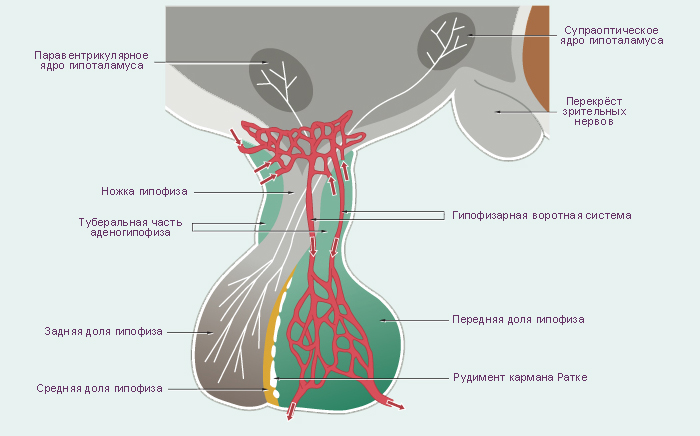
Схема. Гипофиз и окружающие его структуры. Кавернозный синус. Срединное фронтальное сечение мозга человека.
Модификация: Gray H., (1821–1865), Drake R., Vogl W., Mitchell A., Eds. Gray's Anatomy for Students. Churchill Livingstone, 2007, 1150 p., см.: Анатомия человека: Литература. Иллюстрации.
|
 |
|
|
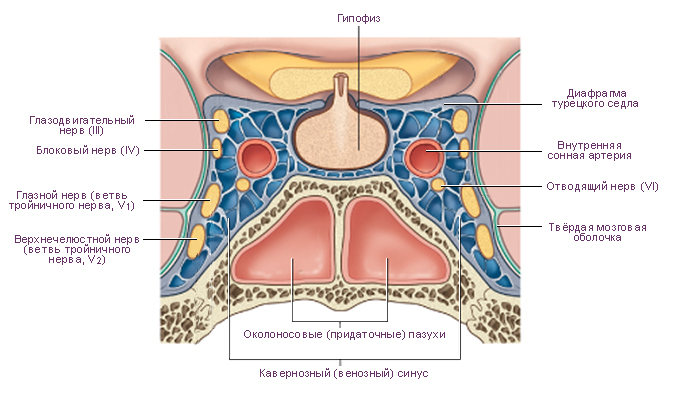
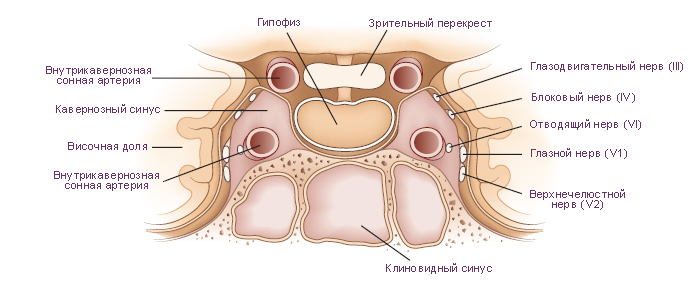
Схема. Поперечное сечение структур в облсти турецкого седла черепа.
Модификация: Melmed S., Polonsky K.S., Larsen P.R., Kronenberg H.M., Eds. Williams Textbook of Endocrinology, 12th ed., Saunders, 2011, 1816 p., см.: Физиология человека: Литература. Иллюстрации.
|
 |
|
Примечание:
|
На верхней (мозговой) поверхности расположено углубление, которое называют турецким седлом. В центре турецкого седла имеется гипофизарная ямка. В ней размещается гипофиз. Латеральные стенки турецкого седла состоят из костной ткани или из ткани твёрдой мозговой оболочки и граничат с кавернозными синусами. Через отверстия в этих стенках проходят парные третий, четвёртый, шестой черепные нервы и внутренняя сонная артерия. Таким образом, при увеличении объёма содержимого области турецкого седла содержимое кавернозных синусов подвержено наибольшему влиянию давления. Крыша, образованная твёрдой мозговой оболочкой, демпфирует колебания давления цереброспинальной жидкости на гипофиз. Непосредственно выше диафрагмы турецкого седла и кпереди от ножки гипофиза расположен перекрест зрительных нервов (зрительный перекрест). Из-за такой топографии, зрительные тракты и соседние центральные структуры являются наиболее чувствительными к повышению давления при увеличении массы гипофиза. Такое давление может поднимать диафрагму турецкого седла, преодолевая её упругое сопротивление. |
|
|
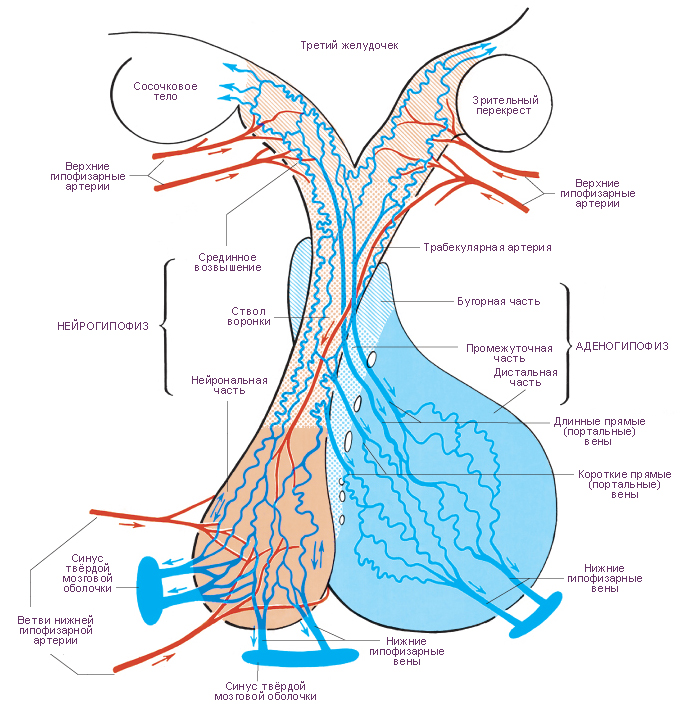
Схема. Кровеносные сосуды гипоталамуса: срединного возвышения, воронки серого бугра; гипофиза и смежных структур.
Модификация: Gray H., (1821–1865), Standring S., Ed. Gray's Anatomy: The Anatomical Basis of Clinical Practice. 39th ed., Churchill Livingstone, 2008, 1600 p., см.: Анатомия человека: Литература. Иллюстрации.
|
 |
|
Примечание:
|
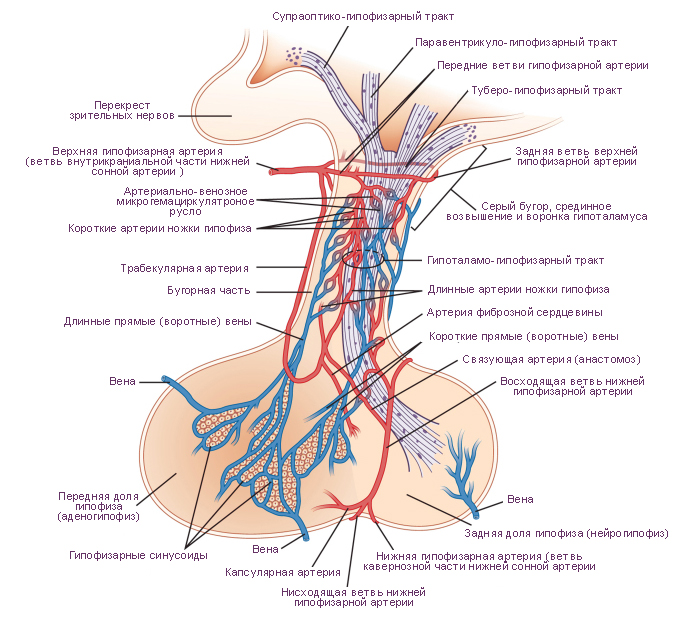
Наиболее вероятная структура гемациркуляторного русла гипоталамуса и гипофиза.
Артерии гипофиза исходят от каждой внутренней сонной артерии по одиночной нижней гипофизарной артерии и по нескольким верхним гипофизарным артериям. Нижняя гипофизарная артерия разделяется на медиальную и латеральную ветви. В нейрогипофизе эти артериальные ветви образуют два типа гемакапиллярных сетей. В первом типе гемакапиллярных сетей приносящими и выносящими сосудами являются артериолы. Такой тип сетей называется артериально-артериальным микрогемациркуляторным руслом. Во втором типе гемакапиллярных сетей нейрогипофиза приносящим сосудом является артериола, а выносящим сосудом является венула. Такой тип сетей называется артериально-венозным микрогемациркуляторным руслом. По венулам артериально-венозного микрогемациркуляторного русла кровь оттекает по трём направлениям. Первое направление потока крови - в нижние гипофизарные вены, а далее в синусы твёрдой оболочки головного мозга. Второе направление потока крови - в длинные и короткие вены и венулы аденогипофиза. Венулы аденогипофиза дают начало его венозноно-венозному микрогемациркуляторному руслу. Выносящими сосудами этой гемакапиллярной сети являются нижние гипофизарные вены аденогипофиза. Третье направление потока венозной крови - от гемакапиллярной сети нижней части воронки (артериально-венозное гемациркуляторное русло). Приносящими сосудами этой сети являются верхние гипофизарные артерии. Кроме того, верхние гипофизарные артерии являются приносящими сосудами, проходящими по оси воронки серого бугра, образующими анастомозы с выносящими нижними гипофизарными артериями. Эти анастомозы образуют гемакапиллярную сеть (артериально-артериальное гемациркуляторное русло) воронки. Верхние гипофизарные артерии снабжают кровью срединное возвышение и образуют в верхней части воронки гемакапиллярную сеть (артериально-венозное гемациркуляторное русло).
Артерии срединного возвышения и стока воронки завершаются характерной сетью капилляров, которая наиболее сложна в верхней части воронки. В срединном возвышении капилляры образуют наружное, «покровное» сплетение капилляров и внутреннее, «глубокое» сплетение капилляров. В гемакапилляры наружного сплетения кровь поступает от верхних гипофизарных артерий, а оттекает по длинным прямым венам. Прямые вены нисходят в переднюю долю гипофиза (аденогипофиз). Внутреннее гемакапиллярное сплетение образовано параллельными цепями наружного гемакапиллярного сплетения. От капилляров нижней части воронки, от её стока кровь оттекает по коротким прямым венам в переднюю долю гипофиза. Как длинные, так и короткие прямые вены впадают в венозные синусоиды, расположенные в тяжах, разделяющих островки секреторных клеток аденогипофиза. Аденогипофиз не снабжается непосредственно артериями. Поскольку как приносящие гемасосуды, так и выносящие гемасосуды аденогипофиза являются однотипными, в частности венами, гемациркуляторное русло аденогипофиза по определению является воротным (портальным) гемациркуляторным руслом, а кровообращение по этим сосудам - воротной (портальной) системой гемациркуляции. Воротная система гемациркуляции предназначена для транспорта либеринов и статинов гипоталамуса. Эти гормоны синтезируются и выводятся мелкими (парвоцеллюлярными) нейросекреторными клетками, сгруппироваными в гипоталамусе. Гипоталамические либерины и статины управляют секреторными циклами аденогипофиза. Промежуточная часть гипофиза не имеет чётко организованного кровоснабжения.
Существует три возможных пути венозного кровотока в гипофизе. Первый путь: приток по длинным и коротким прямым (портальным, воротным) венам. Второй путь: отток по крупным нижним гипофизарным венам в венозные синусы твёрдой мозговой оболочки. Третий путь: отток к гипоталамусу от сети капилляров, расположенных в срединном возвышении. Венозный кровоток предназначен для переноса гормонов от гипофиза к органам-мишеням и клеткам-мишеням. Эти гормоны являются средством управления секрецией (положительная обратная связь). Следует иметь в виду, что гемациркуляция в гипофизе не является полностью изолированной от системного гемациркуляторного русла. Небольшое число вен, участвующих в кровообращении гипофиза, непосредственно связано с венами системного гемациркуляторного русла.
|
|
|
Схема. Гемациркуляторное русло гипоталамуса и гипофиза. Сагиттальное сечение мозга. Вид слева.
Модификация: Melmed S., Polonsky K.S., Larsen P.R., Kronenberg H.M., Eds. Williams Textbook of Endocrinology, 12th ed., Saunders, 2011, 1816 p., см.: Физиология человека: Литература. Иллюстрации.
|
 |
|
Примечание:
|
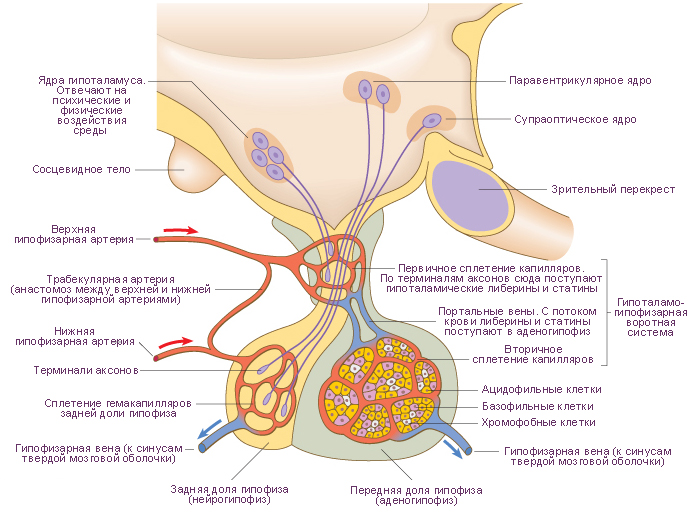
Гипофиз - это эндокринная железа, состоящая из трёх частей: передняя доля гипофиза (~70 ÷ 80 % всей массы гипофиза), задняя доля гипофиза и промежуточная (рудиментарная) доля гипофиза. Гипофиз расположен в гипофизарной ямке турецкого седла клиновидной кости черепа. Железа отделена от полости черепа отростком твёрдой оболочки головного мозга, образующим диафрагму седла. Через отверстие в этой диафрагме гипофиз посредством ножки соединен с вышележащими отделами головного мозга: с воронкой гипоталамуса, с медианным возвышением гипоталамуса промежуточного мозга. Поперечный размер гипофиза ~10 ÷ 17 мм, переднезадний - ~5 ÷ 15 ÷ мм, вертикальный ~5 ÷ 10 мм. Масса гипофиза (~400 ÷ 900 г). У мужчин среднее значение массы гипофиза составляет ~0,5 г, у женщин - ~0,6 г. Поперечное среднее значение диаметра гипофиза составляет ~13 мм. Вертикальный размер - ~6 ÷ 9 мм. Переднезадний размер - ~9 мм. Эти размеры могут изменяться у женщин во время менструального цикла и различаться у неоднократно рожавших женщин. Во время беременности эти размеры могут увеличиваться во всех направлениях. Соответственно увеличивается масса гипофиза до ~1 г. Снаружи гипофиз покрыт капсулой.
Передняя доля гипофиза, или аденогипофиз более плотная, чем задняя доля, нейрогипофиз. В передней доле выделяют три части. Дистальная (передняя) часть аденогипофиза занимает переднюю область гипофизарной ямки турецкого седла черепа. Промежуточная часть аденогипофиза расположена на границе с задней долей гипофиза. Бугорная (туберальная) часть аденогипофиза расположена выше и соединяется с воронкой и медианным возвышением гипоталамуса.
В связи с особенностями кровоснабжения частей гипофиза, передняя доля имеет бледно-желтый цвет, с красноватым оттенком. Паренхима передней доли гипофиза представлена несколькими типами железистых клеток, расположенных скоплениями (трабекулами или фолликулами) разделёнными перегородками из рыхлой соединительной ткани. В толще перегородок располагаются синусоидальные кровеносные капилляры.
Задняя доля гипофиза (неирогипофиз) состоит из нейрональной части и воронки. Нейрональная часть находится в задней области гипофизарной ямки, располагающейся позади бугорной части аденогипофиза. Нейрогипофиз содержит нейроглиальные клетки (питуициты), нервные волокна, идущие в нейрогипофиз от нейросекреторных ядер гипоталамуса, и нейросекреторные тельца. |
|
|
Схема. Главные средства управления эндокринной секреторной активностью гипофиза.
Модификация: Gray H., (1821–1865), Standring S., Ed. Gray's Anatomy: The Anatomical Basis of Clinical Practice. 39th ed., Churchill Livingstone, 2008, 1600 p., см.: Анатомия человека: Литература. Иллюстрации.
|
 |
|
Примечание:
|
В гипофизе выделяют две доли: передняя доля гипофиза (аденогипофиз) и задняя доля гипофиза (нейрогипофиз).
1. Передняя доля гипофиза. Переднюю долю гипофиза разделяют на три части: (а) передняя часть аденогипофиза, (б) промежуточная часть аденогипофиза, (в) туберальная часть аденогипофиза.
(а) Передняя часть аденогипофиза образована скоплениями эпителиальных клеток, образующих разветвленные тяжи - трабекулы, а также нерегулярные фолликулы. В промежутках между скоплениями клеток расположены синусоидальные кровеносные капилляры. Тонкие стенки синусоидов образованы рыхлой волокнистой соединительной тканью. Трабекулы и фолликулы содержат железистые клетки - эндокриноциты.
Различают три типа эндокриноцитов трабекул. Эндокриноциты, располагающиеся по периферии клеточных скоплений, содержат в своей цитоплазме секреторные гранулы, которые легко окрашиваются. В связи с этим такие клетки именуются хромофильными эндокриноцитами. Эндокриноциты, занимающие середину клеточного скопления, имеют нечеткие границы, поскольку их цитоплазма слабо окрашивается. Такие клетки именуются хромофобными эндокриноцитами.
Секреторные гранулы хромофильных эндокриноцитов могут окрашиваться либо основными, либо кислыми красителями. В соответствии с этим хромофильные эндокриноциты разделяют на базофильные и ацидофильные. Гранулы эндокриноцитов содержат гликопротеиды, являющиеся материалом для биосинтеза гормонов. Сравнительно крупные базофильные эндокриноциты в норме составляют ~4 ÷ 10 % общего количества аденоцитов. Выделяют две разновидности базофильных эндокриноцитов.
Клетки первой разновидности характеризуются округлой или овальной формой и эксцентрическим положением ядер. Их секреторные гранулы имеют диаметр ~200 ÷ 300 нм. Количество базофильных эндокриноцитов увеличивается при интенсивной выработке гонадотропных гормонов (гонадотропинов). В связи с этим такие клетки называются гонадотропоцитами, или гонадотропными эндокриноцитами. Одни из гонадотропоцитов вырабатывают фолликулостимулирующий гормон (фоллитропин), а другие - лютеинизирующий гормон (лютропин). Фоллитропин является средством управления формированием половых клеток. Лютропин является стимулирующим средством управления образованим жёлтого тела в яичнике, а также средством управления выработкой мужского полового гормона интерстициальными клетками яичка.
Базофильные клетки второй разновидности отличаются неправильной формой. Их секреторные гранулы очень мелкие (диаметр ~80 ÷ 150 нм) и легко окрашиваются альдегид-фуксином. Эти гранулы содержат меньше гликопротеинов, чем гранулы гонадотропоцитов. Клетки второй разновидности вырабатывают тиротропный гормон - тиротропин. Тиротропин является стимулирующим средством управления функциями фолликулярных эндокриноцитов щитовидной железы. Отсюда, клетки секретирующие тиротропин называются тиротропоцитами, или тиротропными эндокриноцитами.
Ацидофильные эндокриноциты содержат крупные плотные белковые гранулы, которые легко окрашиваются кислыми красителями. По размерам эти клетки несколько меньше базофильных эндокриноцитов. Их количество составляет ~30 ÷ 35 % всех аденоцитов передней доли гипофиза. Они имеют округлую или овальную форму. Ядра базофильных эндокриноцитов располагаются в центре клетки. У них хорошо развита гранулярная эндоплазматическая сеть. Ацидофильные эндокриноциты также имеют две разновидности. Первая разновидность ацидофильных эндокриноцитов - соматотропоциты, или соматотропные эндокриноциты. Одни соматотропоциты вырабатывают гормон роста, или соматотропин. Соматотропин является средством управления ростом организма. Другие соматотропоциты вырабатывают лактотропный гормон, или пролактин и потому их называют соответственно маммотропоцитами, или маммотропными эндокриноцитами и пролактиноцитами, или пролактотропными эндокриноцитами. Лактотропный гормон (пролактин) является средством управления биосинтезом молока в молочной железе. Интенсивность секреции пролактина усиливается у рожени́ц после родов, во время лактации и вскармливания новорожденного. Кроме того, пролактин продлевает существование и функции желтого тела в яичнике. Секреторные гранулы соматотропоцитов имеют шаровидную форму. Их диаметр составляет ~350 ÷ 400 нм. Маммотропоциты отличаются еще более крупными гранулами овальной или удлиненной формы (длина ~500 ÷ 600 нм, ширина ~100 ÷ 120 нм).
Хромофильные эндокриноциты включают ещё одну группу клеток, расположенных главным образом в центральной части передней доли гипофиза. Эти клетки называют кортикотропными эндокриноцитами, или кортикотропоцитами. Они вырабатывают адренокортикотропный гормон (АКТГ, или кортикотропин). Адренокортикотропный гормон является стимулирующим средством управления секрецией гормонов клетками пучковой зоны коркового вещества надпочечников. Кортикотропоциты имеют неправильную форму, дольчатые ядра, хорошо развитую эндоплазматическую сеть. Их секреторные гранулы имеют строение пузырьков, содержащих в своей полости плотную белковую сердцевину. Между мембраной пузырька и его сердцевиной остается светлое пространство.
Хромофобные клетки составляют ~60 % общего числа аденоцитов. Цитоплазма хромофобных эндокриноцитов слабо окрашивается. В ней не видны секреторные гранулы. Хромофобные клетки имеют разную степень развития и выполняют различные функции. Среди них существуют клетки, уже начавшие специализироваться в базофильные или ацидофильные клетки, но еще не успевшие накопить специфические секреторные гранулы. Другие же, хромофобные клетки, наоборот, являются вполне специализированными, но лишившимися своих базофильных или ацидофильных секреторных гранул вследствие интенсивной или длительной секреции. Небольшое количество неспециализированных хромофобных клеток относят к резервным клеткам.
Среди хромофобных клеток встречаются звездчатые (фолликулозвездчатые) клетки. Это клетки небольшого размера, имеющие длинные ветвистые отростки, посредством которых они соединяются в широкопетлистую сеть. Некоторые из отростков проходят между соседними эндокриноцитами и заканчиваются на стенках синусоидных капилляров. Иногда звездчатые клетки группируются в небольшие фолликулы. В полостях этих фолликулов накапливается гликопротеидный секрет. На апикальных поверхностях таких звездчатых клеток (со стороны просвета фолликула) имеются микроворсинки.
Функции аденогипофиза управляются посредством пептидов и аминов, секретируемых нейронами гипоталамуса. Эти нейросекреторные клетки расположены главным образом в медиальной зоне гипоталамуса, в его дугообразном ядре, в медиальной парвоцеллюлярной части паравентрикулярного и перивентрикулярного ядер.
(б) Средняя (промежуточная) часть аденогипофиза представляет собой узкую полоску эпителия. Эндокриноциты средней части способны вырабатывать белковый или слизистый секрет, который, накапливаясь между соседними клетками, приводит к формированию в средней части аденогипофиза фолликулоподобных кист. От задней части аденогипофиза эпителий средней части отделяется тонкой прослойкой рыхлой соединительной ткани.
Эндокриноциты средней части аденогипофиза вырабатывают меланоцитостимулирующий гормон (меланоцитотропин), а также липотропин - гормон, являющийся средством управления метаболизмом липидов.
(в) Туберальная часть аденогипофиза прилежит к ножке гипофиза и соприкасается с нижней поверхностью медианного возвышения гипоталамуса. Туберальная часть образована эпителиальными тяжами, состоящими из кубических клеток с умеренно базофильной цитоплазмой. От туберальных тяжей в переднюю часть аденогипофиза, так же как от эпителия средней части, отходят трабекулы. В некоторых клетках туберальных тяжей встречаются базофильные гранулы.
Кровоснабжение гипоталамуса и гипофиза выполняет функцию реализации управления аденогипофизарного гормонопоэза.
2. Задняя доля гипофиза (нейрогипофиз). Задняя доля гипофиза содержит в основном клетки эпендимы. Они имеют отростчатую или веретеновидную форму и называются питуицитами задней доли гипофиза. Их многочисленные тонкие отростки заканчиваются в адвентиции кровеносных сосудов или на базальной мембране гемакапилляров.
В задней доле гипофиза накапливаются антидиуретический гормон (вазопрессин) и окситоцин. Гормоны вазопрессин и окситоцин вырабатываются крупными пептидохолинергическими нейросекреторными клетками переднего гипоталамуса. Вазопрессин является средством управления реабсорбцией в мочевых канальцах почки. Окситоцин является средством управления сокращением мускулатуры матки. Аксоны нейросекреторных клеток переднего гипоталамуса собираются в гипоталамо-нейрогипофизарные пучки, входят в заднюю долю гипофиза, где заканчиваются крупными терминалями (тельца Херринга, или накопительные тельца), на стенках гемакапилляров.
Percy Theodore Herring (1872-1967), физиолог, врач, Великобритания. |
|
|
Схема. Трехуровневый механизм управления секреции аденогипофиза.
Модификация: Melmed S., Polonsky K.S., Larsen P.R., Kronenberg H.M., Eds. Williams Textbook of Endocrinology, 12th ed., Saunders, 2011, 1816 p., см.: Физиология человека: Литература. Иллюстрации.
|
 |
|
Примечание:
|
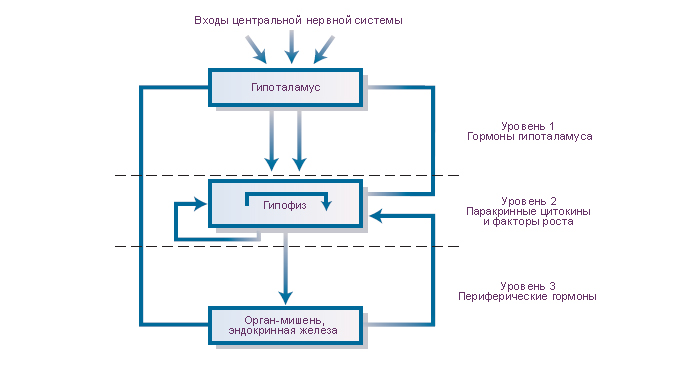
В механизмах управления секрецией передней доли гипофиза можно выделить три уровня. (1) Средством управления секрецией аденогипофиза являются гипоталамические гормоны. Они выводятся в кровь гипоталамо-гипофизарной воротной системы, доставляются в переднюю долю гипофиза и взаимодействуют с биохимическими рецепторами плазмалемм его секреторных клеток. Эти биохимические рецепторы высоко специфичны каждому гипоталамическому гормону. Гипоталамические гормоны передают либо стимулирующие, либо тормозные управляющие сигналы для секреции и выведения гипофизарных тропных гормонов. (2) В модуляции секреции гипофизарных тропных гормонов участвуют (специфическая отрицательная обратная связь) гормоны периферических эндокринных желёз. (3) Гипофизарные паракринные и аутокринные факторы роста и цитокины являются средствами локального регулирования структуры и функций соседних клеток.
Общим результатом реализации этих трёх механизмов является секреция квантов шести гипофизарных тропных гормонов: адренокортикотропный гормон, гормон роста, пролактин, тиреотропный гормон, фолликулстимулирующий гормон и лютеинизирующий гормон. Тропные гормоны выводятся с кровью через кавернозные синусы, каменистые вены, верхнюю полую вену в системную гемациркуляцию. Управление секрецией гормонов гипофиза критически важно для интеграции функций периферических эндокринных желёз, которые, в свою очередь являются средством гуморального управления многими функциями организма. |
|
|
Схема. Развитие передней доли гипофиза = Model for development of the human anterior pituitary gland.
Модификация: Melmed S., Polonsky K.S., Larsen P.R., Kronenberg H.M., Eds. Williams Textbook of Endocrinology, 12th ed., Saunders, 2011, 1816 p., см.: Физиология человека: Литература. Иллюстрации.
|
 |
|
Примечание:
|
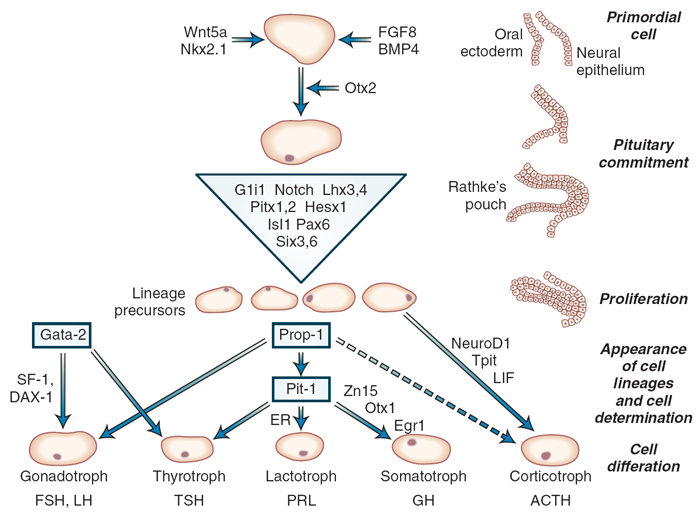
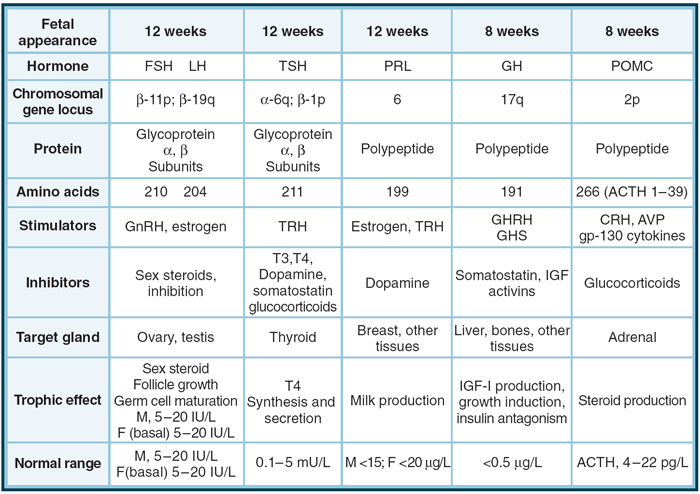
Figure 8-2 Model for development of the human anterior pituitary gland and cell lineage determination by a cascade of transcription factors. Trophic cells
are depicted with transcription factors known to determine cell-specific human or murine gene expression. ACTH , corticotropin; AV P, arginine vasopressin;
CRH, corticotropin-releasing hormone; ER, estrogen receptor; F, female; FSH, follicle-stimulating hormone; GH, growth hormone; GHRH, growth hormone–
releasing hormone; GHS, growth hormone secretagogue; GnRH, gonadotropin-releasing hormone; IGF, insulin-like growth factor; LH, luteinizing hormone; M,
male; POMC, pro-opiomelanocortin; PRL, prolactin; T3, triiodothyronine; T4, thyroxine; TRH, thyrotropin-releasing hormone; TSH, thyrotropin. (Adapted from
Shimon I, Melmed S. Anterior Pituitary Hormones. In: Conn P, Melmed S, eds. Scientific Basis of Endocrinology. Totowa, NJ: Humana Press, 1996; and Amselem S.
Perspectives on the molecular basis of developmental defects in the human pituitary region. In Rappaport R, Amselem S, eds. Hypothalamic-Pituitary Development.
Basel, Switzerland: Karger; 2001; and Dasen JS, O’Connell SM, Flynn SE, et al. Reciprocal interactions of Pit1 and GATA2 mediate signaling gradient–induced determination
of pituitary cell types. Cell. 1999;97:587-598.) с. 187 (178), 2_120 = Williams Textbook of Endocrinology_12ed_2011 |
|
|
Схема. Развитие передней доли гипофиза = Model for development of the human anterior pituitary gland.
Модификация: Melmed S., Polonsky K.S., Larsen P.R., Kronenberg H.M., Eds. Williams Textbook of Endocrinology, 12th ed., Saunders, 2011, 1816 p., см.: Физиология человека: Литература. Иллюстрации.
|
 |
|
Примечание:
|
Figure 8-2 Model for development of the human anterior pituitary gland and cell lineage determination by a cascade of transcription factors. Trophic cells
are depicted with transcription factors known to determine cell-specific human or murine gene expression. ACTH , corticotropin; AV P, arginine vasopressin;
CRH, corticotropin-releasing hormone; ER, estrogen receptor; F, female; FSH, follicle-stimulating hormone; GH, growth hormone; GHRH, growth hormone–
releasing hormone; GHS, growth hormone secretagogue; GnRH, gonadotropin-releasing hormone; IGF, insulin-like growth factor; LH, luteinizing hormone; M,
male; POMC, pro-opiomelanocortin; PRL, prolactin; T3, triiodothyronine; T4, thyroxine; TRH, thyrotropin-releasing hormone; TSH, thyrotropin. (Adapted from
Shimon I, Melmed S. Anterior Pituitary Hormones. In: Conn P, Melmed S, eds. Scientific Basis of Endocrinology. Totowa, NJ: Humana Press, 1996; and Amselem S.
Perspectives on the molecular basis of developmental defects in the human pituitary region. In Rappaport R, Amselem S, eds. Hypothalamic-Pituitary Development.
Basel, Switzerland: Karger; 2001; and Dasen JS, O’Connell SM, Flynn SE, et al. Reciprocal interactions of Pit1 and GATA2 mediate signaling gradient–induced determination
of pituitary cell types. Cell. 1999;97:587-598.) с. 187 (178), 2_120 = Williams Textbook of Endocrinology_12ed_2011 |
|
|
Схема. Действие гормона роста = Action of growth hormone.
Модификация: Melmed S., Polonsky K.S., Larsen P.R., Kronenberg H.M., Eds. Williams Textbook of Endocrinology, 12th ed., Saunders, 2011, 1816 p., см.: Физиология человека: Литература. Иллюстрации.
|
 |
|
Примечание:
|
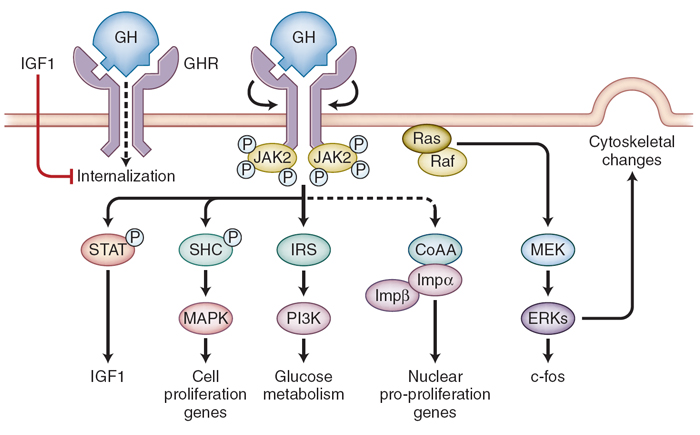
Figure 8-16 Action of growth hormone (GH). GH binds to the growth hormone receptor (GHR) dimer, which undergoes internal rotation, resulting in Jak2
phosphorylation (P) and subsequent signal transduction. Ligand binding to a preformed GHR dimer results in internal rotation and subsequent phosphorylation
cascades. GH signaling is mediated by Jak2 phosphorylation of depicted signaling including Src/ERK pathways. STAT , SHC, and IRS cascades. GH targets
include insulin-like growth factor 1 (IGF1), c-fos, cell proliferation genes, glucose metabolism, and cytoskeletal proteins. GHR internalization and translocation
(dotted lines) induce nuclear proproliferation genes via importin б/в (Impб/Impв) coactivator (CoAA) signaling. IGF1 may also block GHR internalization,
acting in a feedback loop. ERK, extracellular signal-related kinase; IRS, insulin receptor substrate; JAK 2, Janus kinase 2; MEK, dual specifying mitogen-activated
kinase 2. (From Lupu F, Terwilliger JD, Lee K, et al. Roles of growth hormone and insulin-like growth factor 1 in mouse postnatal growth. Dev Biol. 2001;229:141;
and Melmed S. Acromegaly pathogenesis and treatment. J Clin Invest. 2009;119:3189-3202.) |
|
Таблица. Оценка функций аденогипофиза. c. 218.
Модификация: Melmed S., Polonsky K.S., Larsen P.R., Kronenberg H.M., Eds. Williams Textbook of Endocrinology, 12th ed., Saunders, 2011, 1816 p., см.: Физиология человека: Литература. Иллюстрации.
|
| № |
Test* |
Dose |
Normal Response |
Side Effects |
| Адренокортикотропный гормон (адренокортикотропин, кортикотропин); adrenocorticotropic hormone, ACTH |
| 1 |
Insulin tolerance |
0.1-0.15 U/kg IV |
Peak cortisol response >18 μg/dL, or ↑
≥ 5 μg/dL |
Sweating, palpitation, tremor |
| 2 |
Metyrapone |
30 mg/kg PO at 11 p.m. |
Peak 11-DOC ≥ 7 μg/dL
Peak cortisol ≤ 7 μg/dL
Peak ACTH >75 pg/mL |
Nausea, insomnia, adrenal crisis |
| 3 |
CRH stimulation |
100 μg IV |
Peak ACTH ≥ 2-4-fold
Peak cortisol ≥ 20 μg/dL or ↑ ≥ 7 μg/dL |
Flushing |
| 4 |
ACTH stimulation |
250 μg IV or IM, or 1 μg IV |
Peak cortisol ≥ 20 μg/dL |
Rare |
| Тиролиберин; thyroid-stimulating hormone, TSH |
| 5 |
Serum T4 (free T4)
Total T3
TSH—third generation
TRH stimulation |
200-500 μg IV |
Peak TSH ≥ 2.5-fold, or ↑ ≥ 5-6 mU/L
(females) or ≥ 2-3 mU/L (males) |
Flushing, nausea, urge to micturate |
| Пролактин; prolactin PRL |
| 6 |
Serum PRL
TRH stimulation |
200-500 μg IV |
PRL ↑ ≥ 2.5-fold |
Flushing, nausea, urge to micturate |
| Лютеинизирующий гормон / фолликулстимулирующий гормон; LH, luteinizing hormone / FSH, follicle-stimulating hormone |
| 7 |
Serum LH and FSH
Serum testosterone
GnRH Stimulation |
100 μg IV |
Elevated in menopause and in men with
primary testicular failure (otherwise
normal) 300-900 ng/mL
LH ≥ 2-3-fold, or ↑ by 10 IU/L
FSH ≥ 1.5-2-fold, or ↑ ≥ 2 IU/L |
Rare |
| Гормон роста; growth hormone, GH |
| 8 |
Insulin tolerance |
0.1-0.15 U/kg |
GH peak >5 мg/L |
Sweating, palpitation, tremor |
| 9 |
L-Arginine Arginine |
0.5 g/kg (maximum,
30 g) IV over 30-120 min |
GH peak >0.4 мg/L |
Nausea |
| plus |
| 10 |
GHRH |
GHRH 1-5 мg/kg |
GH peak >4 мg/L |
Flushing |
|
Обозначения:
|
ACTH, adrenocorticotropic hormone;
CRH, corticotropin-releasing hormone;
11-DOC, 11-deoxycorticosterone;
FSH, follicle-stimulating hormone;
GH, growth hormone;
GHRH, growth hormone–releasing hormone;
GnRH, gonadotropin-releasing hormone; |
LH, luteinizing hormone;
PRL, prolactin;
T3, triiodothyronine;
T4, thyroxine;
TSH, thyroid-stimulating hormone;
TRH, thyrotropin-releasing hormone. |
|
|
Схема. Влияние изменений осмоляльности плазмы крови на концентрацию аргинин-вазопрессина в плазме и последующее влияние на осмоляльность мочи и объём мочевыведения = Effect of change in plasma osmolality (pOsm, in mOsm/kg of
H2O) on plasma arginine vasopressin (PAVP, in pg/mL) and consequent effects
on urine osmolality (UOsm, in mOsm/kg of H2O) and urine volume (L/day)..
Модификация: Melmed S., Polonsky K.S., Larsen P.R., Kronenberg H.M., Eds. Williams Textbook of Endocrinology, 12th ed., Saunders, 2011, 1816 p., см.: Физиология человека: Литература. Иллюстрации.
Примечание:
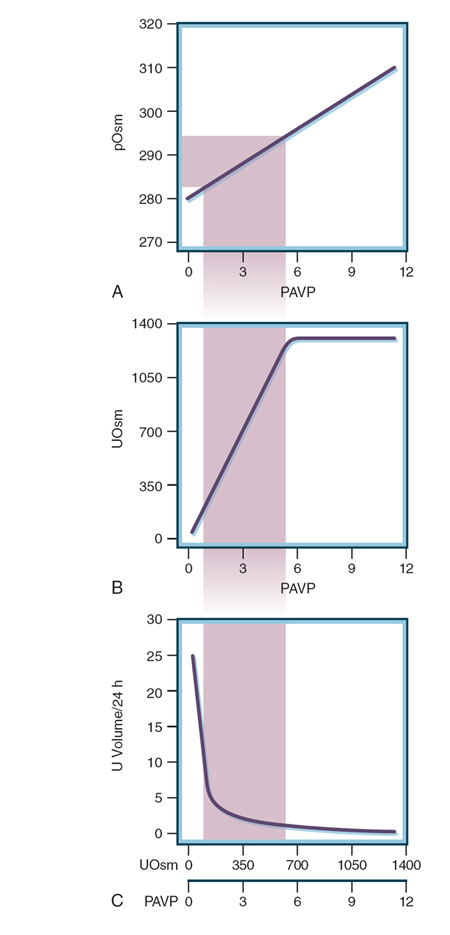
Figure 10-3 Effect of change in plasma osmolality (pOsm, in mOsm/kg of
H2O) on plasma arginine vasopressin (PAVP, in pg/mL) and consequent effects
on urine osmolality (UOsm, in mOsm/kg of H2O) and urine volume (L/day).
The shaded area represents the normal range. A, Small changes in pOsm
induce changes in PAVP, typically between less than 0.5 and 5 to 6 pg/mL.
B, Changes in PVAP induce changes in UOsm through the full range, from
maximally dilute to maximally concentrated urine. Although PAVP can rise
to higher levels than 6 pg/mL, this does not translate into increased UOsm,
which has a maximum determined by the osmolality of an inner medulla of
the kidney. C, The relationship of urine volume to UOsm is logarithmic,
assuming a constant osmolar load and the urine volume that would excrete
that osmolar load at the UOsm indicated. As a result, urine volume changes
relatively little with small changes in the other parameters until there is
almost complete absence of PAVP, after which the urine volume increases
dramatically. (Calculated from a formula presented in Robertson G, Shelton R,
Athar S. The osmoregulation of vasopressin. Kidney Int. 1976;10:25-37. Figure
by A. G. Robinson, University of Los Angeles, CA, with permission of Macmillan
Publishers, Ltd.) 303 (294) 2_120 |
 |
|
ЭНДОКРИНОЛОГИЯ: ОГЛАВЛЕНИЕ
ЭНДОКРИНОЛОГИЯ: ИЛЛЮСТРАЦИИ.
ЭНДОКРИНОЛОГИЯ: ТАБЛИЦЫ.
ЭНДОКРИНОЛОГИЯ: ЛИТЕРАТУРА.
|
«Я У Ч Е Н Ы Й И Л И . . . Н Е Д О У Ч К А ?»
Т Е С Т В А Ш Е Г О И Н Т Е Л Л Е К Т А
Предпосылка:
Эффективность развития любой отрасли знаний определяется степенью соответствия методологии познания - познаваемой сущности.
Реальность:
Живые структуры от биохимического и субклеточного уровня, до целого организма являются вероятностными структурами. Функции вероятностных структур являются вероятностными функциями.
Необходимое условие:
Эффективное исследование вероятностных структур и функций должно основываться на вероятностной методологии (Трифонов Е.В., 1978,..., ..., 2015, …).
Критерий: Степень развития морфологии, физиологии, психологии человека и медицины, объём индивидуальных и социальных знаний в этих областях определяется степенью использования вероятностной методологии.
Актуальные знания: В соответствии с предпосылкой, реальностью, необходимым условием и критерием...
...
о ц е н и т е с а м о с т о я т е л ь н о:
— с т е п е н ь р а з в и т и я с о в р е м е н н о й н а у к и,
— о б ъ е м В а ш и х з н а н и й и
— В а ш и н т е л л е к т !
|
♥ Ошибка? Щелкни здесь и исправь ее! Поиск на сайте E-mail автора (author): tryphonov@yandex.ru
|